|
Путеводитель по Крыму
Группа ВКонтакте:
Интересные факты о Крыме:
В Крыму действует более трех десятков музеев. В числе прочих — единственный в мире музей маринистского искусства — Феодосийская картинная галерея им. И. К. Айвазовского. На правах рекламы: • Кладочная сетка купить сетку на сайте. |
Главная страница » Библиотека » И.И. Пузанов. «Крым: Путеводитель»
И.И. Пузанов. «Черное море»Посвящается памяти Н.И. Андрусова Я видел море: я его
Полежаев. Чувства, которые испытывал поэт, попав на берег моря конечно, и теперь испытывает всякий чуткий к природе человек. Чувства эти сводятся, во-первых, к восторженному преклонению перед величием морского простора, во-вторых, к жадному стремлению постигнуть его тайны. Однако, во всю первую половину 19-го века, соответственно слабому развитию у нас науки, эмоциональная сторона в восприятии Черного моря явно преобладала; лишь отдельные ученые — Паллас, Ратке, Нордманн, делали попытки к изучению моря, главным образом его животного населения. Лишь начиная с 60-х годов, соответственно быстрому развитию океанографической науки на Западе, начинается и у нас систематическое изучение Черного моря. Из числа научных экспедиций, работавших над изучением Черного моря, следует особенно отметить изыскания военного моряка С.О. Макарова в области Босфора, в 1886 г., знаменитые «глубокомерные экспедиции» на судах военного флота, имевшие место в 1890—91 гг. с участием Н.И. Андрусона, И.П. Шпиндлера, А.А. Остроумова и др., и многочисленные поездки бывшего заведующего Севастопольской биостанцией С.А. Зернова, совершенные между 1902 и 1914 годами. После продолжительного затишья в деле научного изучения Черного моря, вызванного войной, работа с новой силой возобновляется только, примерно, с 1922 г. Главным управлением рыболовства в Москве организуется большая «Азовско-черноморская научно-промысловая экспедиция», под руководством авторитетнейшего исследователя русских морей, профессора Н.М. Книповича. Энергично возобновляет свою деятельность и Севастопольская биостанция, которая работает рука об руку с Главным гидрографическим правлением и Убекочерназом,1 используя для своих экспедиций, помимо собственных судов, и крупные суда Черноморского военного флота. В настоящее время, помимо Севастополя, существуют биостанции в Варне, Одессе, Карадаге и Новороссийске. Дружные исследования самых последних лет значительно расширили и углубили наше знакомство с водным режимом и биологией Черного моря, и данные этих исследований широко использованы при составлении настоящего очерка. В изучении геологического прошлого Черного моря, которое одно может дать нам ключ к пониманию его настоящего, руководящую роль играли многочисленные работы покойного академика Н.И. Андрусова, организатора и участника классических «глубокомерных» экспедиций 1890-х годов, и одного из лучших знатоков Черного моря в его прошлом и настоящем. Незабвенной памяти этого неутомимого исследователя мы и посвящаем этот очерк. В настоящее время Черное море несет это название на всех европейских языках: Black Sea англичан, Schwarzes Meer немцев, Mer Noire французов, Кара-Дениз турок и татар, Маври Таласса греков. Название это обязано своим происхождением темному оттенку морской синевы, явственно ощущаемому мореплавателем, попадающим в Черное море непосредственно из Эгейского, именуемого иногда современными греками, по контрасту, «аспро таласса» (белое море). Но предки современных греков иначе называли наше море: негостеприимным Понтом (Понтос аксинос), когда берега его были населены враждебными варварами, гостеприимным морем (Понтос эвксинос), когда они покрылись цветущими греческими колониями. Черное море со всех сторон окружено сушей и сообщается с Мраморным при помощи узкого пролива — Босфора Фракийского. Особенностями его являются правильность формы, простота подводного рельефа и береговой линии, которая редко нарушается глубокими бухтами и мысами. Исключение составляет северный берег, где полуостров Крым, глубоко вдаваясь к югу, отделяет Одесский и Каркинитский заливы от Азовского моря; последнее есть не более, как глубокий залив Черного, сообщающийся с ним при помощи Керченского пролива (в древности — Босфора Киммерийского). Простота береговой линии сказывается и в отсутствии островов: в сущности, единственными настоящими островами являются о. Фидониси или Змеиный, против устья Дуная, Березань против Очакова, Кефкен близ Анатолийского берега. Наибольшая длина Черного моря с В. на З. — 1160 километров, наименьшая, с С. на Ю. — 270 километров, поверхность его — 411 540 кв. километров. Черное море является одним из очень глубоких внутренних морей: большая часть его огромной, ровной центральной котловины занята пучинами свыше 2000 метров; наибольшая из измеренных глубин — 2248 метров лежит на пересечении Севастопольского меридиана с параллелью Сухума.2 Эта глубокая котловина вдоль берегов опоясана, так называемой, «континентальной ступенью», т. е. подводной террасой, поверх которой лежат глубины менее 180 метров (или 100 морских, т. е. 6-футовых саженей). Обращенный к морю склон континентальной ступени отличается значительной крутизной; профессор Остроумов весьма наглядно сравнивает котловину Черного моря с глубокой тарелкой, края которой соответствуют континентальной ступени. Край континентальной ступени, или иначе линия 180-метровой глубины (так называемая, стосаженная изобата) ближе всего подходит к берегу у южного побережья Крыма, Кавказа и Анатолии; С.-З. часть моря гораздо мельче и занята глубинами ниже 80 метров, Азовское же море — совсем мелководно, и глубина его, в общем, не превышает 13 мет. Черное море издревле славилось своими бурями, которые достигают своей максимальной силы в зимние месяцы. Особенно страшны С.-З. и С.-В. штормы: последние известны под названием «бора» и сопровождаются обычно сильными морозами. Суда, застигнутые «борой», иногда гибнут от тяжести льда, намерзающего на их оснастке. Особенно опасна бора у С.-В. побережья Кавказа; юго-восточный угол Черного моря, защищенный Кавказским хребтом, отличается, наоборот, чрезвычайным спокойствием атмосферы. Волны, вздымаемые на Черном море бурями, конечно, не могут сравниться своею высотой с океанскими, но все же достигают высоты 4-х метров и страшны своею неправильностью и крутизной.3 Разумеется, в таком закрытом со всех сторон бассейне, как Черное море, не могут ощущаться приливы и отливы открытого океана; однако, под влиянием притяжения луны и солнца, обширная водная поверхность нашего моря дает собственные приливы и отливы, максимальная амплитуда которых может достигать по наблюдениям в Севастополе 3,6 сантиметров, что, конечно, не имеет никакого практического значения. Гораздо важнее осеннее понижение морского уровня у северных берегов, происходящее в результате летнего испарения и северных ветров, и достигающее в среднем величины 1 фута, в отдельных случаях — до 3-х футов.4 Течения Черного моря вызываются, главным образом, преобладающими ветрами, и в общем, направлены вдоль его берегов против часовой стрелки. Однако, общепринятая схема двух круговоротов, окружающих западную и восточную области спокойной воды (так называемые, «галистатические» области), и разделенные выступами Крымского и Синопского полуостровов, по последним исследованиям вряд ли соответствует действительности, особенно в отношении восточной части моря. Течения Азовского моря отличаются своим непостоянством, но в общем, имеют такое же круговое направление «против часовой стрелки», как и в Черном море.
С физической стороны воду всякого моря принято характеризовать прежде всего со стороны ее прозрачности и цвета. Вблизи от берегов, на мелководьи, прозрачность морской воды, в том числе и черноморской, невелика, вследствие массы взвешенных в ней твердых частиц, и вода имеет зеленоватый, иногда даже желтоватый оттенок. Но над большими глубинами прозрачность воды Черного моря довольно значительна: окрашенный в белый цвет «диск Секки», диаметром в 50 см, можно еще различить до глубины 20 м, а морская пучина имеет красивый голубой цвет с легким зеленоватым отливом. При большом различии климатических условий в области Черного моря температура поверхностных слоев, естественно, весьма различна в разных местах, особенно зимой. Наивысшая температура наблюдается в августе и может достигать 29,0°Ц у Одессы, Севастополя и Батума, 25° у Ялты. Самые низкие температуры наблюдаются в феврале: в среднем, 1,2 — у Одессы, 0,7 — у Очакова, 5,4 — у Севастополя, 6,0 — у Ялты, 8,3 — у Батума. Так как морская вода соленостью в 1,8% замерзает лишь при охлаждении до −1°Ц, глубокие части Черного моря, конечно, замерзнуть не могут; но в мелководной С.-З. части моря вода у берегов в сильные морозы охлаждается до самого дна, и потому часто покрывается льдом, иногда на большие пространства; так, в необычайно суровую зиму 1911 г. замерзла вся поверхность моря к северу от линии Одесса—Тарханкут. Что касается слабосоленого и мелкого Азовского моря, то частичное замерзание его, равно как и Керченского пролива, повторяется каждую суровую зиму, и рыбаки местами переезжают его на санях. Однако, значительные колебания температуры Черного моря наблюдаются лишь в самых поверхностных его слоях; на глубине ниже 50 метров температурные колебания делаются уже незначительными, не превышая 1°Ц в течение года (от 7,5 в феврале до 8,4 в августе). На глубине 200 м они совсем исчезают, и вода круглый год сохраняет температуру 8,7°. Присутствием на сравнительно небольшой глубине очень холодной воды объясняется общеизвестное явление так называемого «сгона», то есть внезапного захолодания воды летом, при С. и С.-З. ветрах, особенно при условии прохождения глубокого циклона. Если ветер достаточно силен и продолжителен, он отгоняет от берегов сравнительно тонкий слой прогретой воды, которая замещается холодной водой из глубины. По прекращении ветра равновесие скоро восстанавливается, и теплый слой, так сказать, возвращается на свое место. Начиная с глубины 50 метров и ниже, температура, хотя медленно, но неизменно возрастает, достигая у самого дна почти 9° (8,95°)Ц. Весьма важно также отметить, что вышеупомянутая граница постоянной температуры лежит на глубине 200 метров лишь посредине моря, опускаясь у берегов до глубины 300 метров. То же относится и к другим изотермическим поверхностям (т.е. поверхностям одинаковой температуры), которые все имеют выпуклую, куполообразную форму. Возникает вопрос, как объяснить вышеописанное распределение температур, которое кажется парадоксальным, так как ниже 50 м слои более теплые лежат выше слоев более холодных. Парадокс этот только кажущийся, так как распределение различных слоев воды зависит от их плотности, которая сама есть функция не только температуры, но и солености воды, к рассмотрению которой мы сейчас переходим. По сравнению с другими морями вода Черного моря отличается пониженною соленостью. В то время, как в океане соленость воды равна, в среднем 3,5%. Средиземном море — 3,6%, в Черном море поверхностные слои воды содержат, в среднем, всего 1,8% соли. В районе действия больших рек — Днепра, Днестра, особенно Дуная, соленость значительно меньше, падая даже в значительном расстоянии от берега до 1,3%. У берегов Одессы она, в среднем, равна 1,45%, а в Азовском море колеблется от 1,5 до 1,0 и ниже. Но помимо меньшей концентрации солей, вода Черного моря отличается также иным их составом. В океане мы находим в 100 частях сухой, выпаренной морской соли 77,75 частей поваренной соли, 4,74 серно-кислого магния и 3,60 частей гипса; в Черном — 77,72 поваренной соли, 7,11 сернокислого магния и 2,58 гипса; поэтому вода Черного моря является более горькой.5 С глубиной соленость воды Черного моря увеличивается, достигая уже на глубине 50 м 2,0%. Но опять-таки столь быстрое возрастание наблюдается лишь в центре моря; у берегов 2% солености мы встречаем лишь на глубине около 175 м. Иными словами изогалины, т. е. поверхности одинаковой солености, тоже имеют куполообразную форму. На глубине 400 м соленость достигает 2,2%, и такой остается до самых больших глубин. Итак, глубины Черного моря заполнены хотя несколько более теплой, но зато более соленой, и в результате — более плотной водой.6 В этом отношении Черное море резко отличается от Средиземного, вода которого до самых больших глубин сохраняет температуру, близкую к 13°. Но главная особенность Черного моря заключается в том, что вся огромная масса его глубинной воды ниже 180 метров глубины заражена сероводородом. Следы последнего начинаются уже на глубине 150 метров (около 0,5 см³ на литр воды, при температуре 0°Ц и давлении 1 атмосферы, в пробах воды, взятых на средине моря); на глубине 180 м количество его достигает уже 1,0, на глубине 300 м превышает 2,0, а у самого дна достигает 6,5 куб. сант. на литр. Итак, вся огромная масса глубинной воды заражена ядовитым зловонным газом, как мы увидим дальше, почти совершенно исключающим всякую органическую жизнь. В этом отношении Черное море стоит совершенно особняком среди других морских бассейнов; лишь в Каспийском море7 и некоторых норвежских фьордах (глубоких морских заливах) обнаружено нечто подобное, но в гораздо более слабой степени. Факт присутствия на глубинах Черного моря сероводорода был открыт Н.И. Андрусовым в 1890 году во время экспедиции на канонерке «Черноморец», но, несомненно, рыбаки еще лет за 25 до этого знали о присутствии на больших глубинах вонючей грязи, от которой чернеют белужные крючки; и по сию пору рыбаки называют глубокую впадину Черного моря «вонючей канавой». Присутствие сероводорода в Черном море всякий может проверить, опустив на длинной бечевке на глубину 200—300 метров серебряную монету, которая быстро почернеет. В тесной, как мы увидим, причинной связи с присутствием на глубине Черного моря сероводорода стоит замечательный факт отсутствия там кислорода — газа, необходимого для существования организмов. В верхних слоях воды, до глубины 40—50 метров, количество его превышает 7 см³ на литр, т. е. приближается к полной насыщенности при данных условиях температуры и солености. Но ниже этих глубин процент кислорода быстро падает и совсем исчезает несколько ниже появления первых следов сероводорода, т. е. в средине моря на глубине 120—150 м, а у берегов — около 200 м. Следовательно, и в этом случае мы должны отметить весьма замечательный и важный факт, что верхняя граница сероводорода и нижняя кислорода образуют как бы куполообразную поверхность, приподнятую в средине моря. Нам предстоит теперь истолковать изложенные выше особенности Черного моря в отношении распределения температуры, солености и газов. Пониженная соленость поверхностных слоев воды Черного моря легко объясняется огромным количеством более легкой пресной воды, вносимой в море многоводными реками, особенно весною и осенью. Вода эта растекается сверху тонкой опресненной пленкой и вытекает из моря через Босфор, на поверхности которого возникает течение, достигающее скорости до 9 км в час и простирающееся до глубины от 18 до 55 метров. В противовес ему, по законам равновесия жидкостей, начиная от указанной глубины и до дна, в проливе работает более слабое нижне-Босфорское течение, по которому в Черное море втекает более тяжелая, соленая и теплая вода Мраморного моря, заполняющая всю глубокую котловину Понта и мало смешивающаяся с поверхностными слоями. В результате такого двоякого пополнения в Черном море и устанавливается, как стационарное явление, указанное выше расслоение воды по проценту содержащихся в ней солей: сравнительно тонкая — в среднем 150 м пленка более легкой воды с соленостью 1,8—2% ложится поверх более плотной массы с соленостью 2—2,2%, простирающейся до дна. Пленка эта, подобно слою масла на воде, как бы закупоривает морские глубины, препятствуя нормальному перемешиванию поверхностных и глубоких слоев. Эта, так называемая, вертикальная циркуляция, в других морях простирается до самого дна, зимой — вследствие непрерывного опускания на дно охлажденных поверхностных слоев, и в менее сильной степени летом — вследствие аналогичного погружения тех же слоев осолоненных в результате интенсивного испарения на поверхности. В Черном море это перемешивание тоже происходит, но лишь до глубины немногим ниже 150 метров, так как опресненная поверхностная вода ни при каких условиях не может достигнуть плотности, которую имеет более соленая вода глубин ниже 150 м, и следовательно, никогда не может погрузиться ниже этой границы. Но если это так, то и кислород, поглощаемый из воздуха верхними слоями воды, тоже никогда не может проникнуть глубже 150 м. Наличие на глубинах сероводорода является лишь неизбежным следствием вышеописанного гидрологического режима. Отсутствие на глубинах кислорода создает там условия, при которых могут существовать только анаэробные, т. е. обходящиеся без кислорода организмы. Таковыми являются многочисленные сероводородные бактерии, в первую очередь из рода Microspira; констатированные в придонном илу и восстанавливающие сероводород из сернокислых солей.8 Этот важный биохимический процесс возможен лишь при условии отсутствия кислорода, и при наличии органического вещества, которое в данном случае доставляется в виде трупов бесчисленных организмов, непрерывным дождем падающих вглубь из поверхностных населенных слоев. Помимо собственно сероводородных бактерий в толще морской воды живут мириады гнилостных бактерий, производящих гниение органического вещества с выделением сероводорода. Покойный Н.И. Андрусов придавал очень большое значение последнему способу происхождения сероводорода, однако, произведенные за последнее время подсчеты показали, что сероводород органического происхождения играет в режиме Черного моря лишь незначительную роль, образуя менее 1/600 общего количества сероводорода, и что главная масса этого газа образуется все же за счет сернокислых солей морской воды. Но каково бы ни было происхождение сероводорода в Черном море, на глубинах выше 150 м он исчезает, быстро окислясь в присутствии кислорода по формуле: H2S + O = H2O + S.9 Было бы ошибкой предполагать, что зараженность глубин Черного моря сероводородом объясняется наличием сероводородных бактерий лишь в этом море: бактерии эти обнаружены и в илу Мраморного моря, но богатство его вод кислородом парализует их деятельность.
Типичные животные Черного моря (по С.А. Зернову). Объяснение рисунков. 1. Медуза корнерот — Pilema pulmo Haeck. 2. Планктонная диатомовая водоросль хэтоцерос — Chaetoceros. 3. Медуза ушастая аврелия — Aurelia aurita M. Edw. 4. Ночесвет или ноктилюка — Noctihica miliaris Ehrb. 5. Перидиниевая водоросль церациум — Ceratium tripos. 6. Ктенофора или ребровик плейробранхия — Pleurobranchia pileus Fabr. 7. Перидиниевая водоросль гониавлакс — Gonyaulax polyedra Stein. 8. Медуза раткеа — Rathkea blumenbachii Rathke. 9. Медуза сарсия — Sarsia tubulosa Forbs. 10. Бурая водоросль цистозира — Cystoseira barbata L. 11. Немертина — Eunemertes gracilis Johnst. 12. Червь нереида — Nereis diversicolor Müll. 13. Червь пескожил — Arenicola grubii Clap. 14. Гидроид аглаофения — Aglaоphenia pluma Lmc. 15. Губка суберитес — Suberites domuncula, Schmidt. 16. Трубчатый червь — Potamoceros triquetroides Panc. 17. Кремневая губка халихондрия — Halichondria grossa Schmidt. 18. Актиния, морской анемон — Actinia equina L. 19. Зеленая водоросль морской салат — Ulva lactuca L. 20. Амфиура — Amphiura florifera Frbs. 21. Красная водоросль — Phyllophora rubens Grev. 22. Зеленая водоросль энтероморфа — Enteromorpha. Примечание. Рисунки № № 2, 4, 5, 7 изображают собою животных, почти невидимых невооруженным глазом Обратимся теперь к неоднократно подчеркнутому нами факту куполообразного строения воображаемых поверхностей, разделяющих в Черном море слои с одинаковой температурой, одинаковым содержанием солей, кислорода, сероводорода и т. д. — т. е. изотерм, изогалин, изоксиген. (Мы видели, что в центре моря поверхность, разделяющая слои с соленостью выше и ниже 2%, лежит на глубине 50 м, а у берегов — 175 м). Профессор Н.М. Книпович видит причину этого явления в круговых течениях, которые, сообщая поверхностным слоям центробежную силу, как бы прижимают их к берегам и тем увеличивают их толщу. Другие исследователи видную роль в опускании у берегов границ кислорода, сероводороде и т. д. приписывают также тому обстоятельству, что благодаря прибою, сгону и нагону вод под влиянием ветров, вертикальная циркуляция простирается у берегов на большую глубину. Однако, в западной части моря куполообразная форма изо-поверхностей сильно искажается под влиянием больших количеств пресной воды, вливаемой Дунаем и большими русскими реками. Гидрологический режим мелководного Азовского моря коренным образом отличается от Черноморского. Благодаря массе пресной воды, вливаемой Доном и Кубанью, и незначительной глубине Керченского пролива вода Азовского моря гораздо менее солона, чем в Черном. Среднюю соленость ее можно определить в 1%, но под влиянием ветров, нагоняющих то пресную воду из р. Дона, то соленую из Керченского пролива, содержание соли сильно колеблется, особенно в Таганрогском заливе. Летом вода прогревается, конечно, до самого дна, и в штилевую погоду устанавливается известное равновесие в распределении температуры, солености и содержания кислорода. Поверхностные слои, нагретые, например до 26°, с соленостью около 1%, могут ложиться поверх более соленой, прохладной воды черноморского происхождения (с соленостью около 1,5 и температурой около 22°). В зимнее время температура воды часто падает ниже 0°, что влечет за собой замерзание обширных площадей его. Известное сходство с гидрологическими условиями Азовского моря мы находим в лиманах, как в бассейне Черного моря называют большие, большей частью вытянутые озера, лежащие у устьев рек — Днестра, Буга, Кубани, Риона и т. д. Здесь мы тоже наблюдаем пониженную соленость и расслоение воды на более опресненный верхний и более соленый нижний слой, в результате поверхностного течения из реки и придонного — из моря. И здесь гидрологический режим отличается крайним непостоянством, завися от ветров и степени наполнения реки. Сиваш является, по существу лишь громадным, причудливой формы сильно изолированным заливом Азовского моря, сообщающимся с ним при помощи узкого пролива у Геническа. В северной части Сиваша соленость лишь немногим больше Азовской — здесь ловят кефаль и камбалу, последняя даже нерестится. В южной же части соленость достигает 16% и больше. Изложенные выше физико-химические особенности Черного моря определяют характер и распределение организмов — животных и растений, населяющих его толщу. Так как сероводород представляет из себя яд для животных, дышащих кислородом (на воздухе уже 0,5% его убивает млекопитающих), вся огромная толща черноморской воды, лежащая ниже 100—200 метров, по необходимости лишена жизни и представляет из себя настоящее царство смерти. Это относится одинаково как к организмам пелагическим, т. е. населяющим толщу морской воды, так и к бентосу, т. е. животным придонным. Изучая население какого-нибудь водоема, принято животных пелагических разделять, во-первых, на планктон, куда относятся взвешенные в воде организмы как животного, так и растительного происхождения, большею частью микроскопические или полумикроскопические, пассивно переносимые волнами и течениями, и нектон, охватывающий животных, обладающих сильными орудиями плавания и потому более или менее независимых в своем передвижении. Из этих 2-х биологических группировок планктон наиболее непосредственно зависит от вышеизложенных гидрологических условий, почему мы с него и начнем изложение органической жизни Черного моря. Лишь немногие организмы черноморского планктона, главным образом медузы, обладают сколько-нибудь значительными размерами; огромное большинство их невидимо, или еле видимо простым глазом. Для добывания их употребляются конические «планктонные» сетки из частого шелкового сита с металлическим стаканом на конце, опускаемые во время хода корабля на различные глубины. Особое устройство позволяет в любой момент захлопывать сетку, благодаря чему становится возможным облавливать слои воды определенной глубины. Как и следовало ожидать, планктонные организмы населяют лишь свободный от сероводорода верхний слой морской воды; поэтому «граница жизни» проходит в толще морской воды, примерно на глубине 200 м вблизи берегов, и 150 м в средине моря, т. е. тоже имеет уже знакомую нам форму куполообразной поверхности. Однако, этот обитаемый слой населен далеко не равномерно: соответственно резкому падению количества растворенного в воде кислорода ниже глубины 50 м в открытом море и 75 м вблизи от берегов, резко понижается и количество планктонных организмов, и сколько-нибудь густое население их мы встречаем лишь выше этой границы «кислородного скачка».
Морской кот По своему составу планктон разделяется на растительный (фитопланктон) и животный (зоопланктон). Первый составляется из неисчислимого количества микроскопически-малых водорослей — диатомовых, сине-зеленых, перидиней и т. д., число видов которых в Азовском море превышает — 10, а в Черном — 80 (рис. 2, 5). Несмотря на большее видовое разнообразие, количество фитопланктона в Черном море сравнительно невелико; вообще говоря, оно заметно больше у берегов, и сильно возрастает к осени. В Азовском же море количество его сравнительно однообразного фитопланктона — огромно, в результате чего с июня по ноябрь вода его, как говорят, «зацветает», окрашиваясь то в зеленый цвет сине-зелеными водорослями — Cyanophyceae, то в буро-красный цвет диатомеями и перидинеями. Красные полосы от скоплений перидиней наблюдаются, впрочем, и в Черном море. Еще большее разнообразие выказывают организмы животного планктона, отличающиеся как правило хрустально-прозрачным телом и нередко причудливыми формами. Сюда относятся разнообразные инфузории, особенно характерны колокольчиковые — Tirtinnoidea, черви, ветвистоусые, веслоногие и иные рачки, медузы, ребровики, аппендикулярии (рис. 1, 6, 39, 43). К ним присоединяются, особенно вблизи от берегов, неисчислимые личинки придонных животных — червей, мшанок, моллюсков, иглокожих, крабов и усоногих. Многие из этих организмов обладают способностью испускать свет, чем объясняется общеизвестное явление свечения моря, неправильно называемое фосфоресценцией. Весной и в начале лета это свечение вызывается одноклеточным жгутиковым организмом ночесветкой — Noctiluca miliaris (р. 4), в конце лета, когда оно особенно эффектно — преимущественно перидиниевыми водорослями — Prorocentrum micans и др. Наиболее крупными, а потому и общеизвестными животными черноморского планктона являются медузы, из коих чаще всего прибивается к берегу аврелия или «морское сердце» — Aurelia aurita (р. 3) — медуза, обладающая лишь слабыми собственными движениями. Другая, гораздо более крупная медуза-корнерот — Pilema pulmo (р. 1), встречающаяся по преимуществу у западных берегов Крыма, довольно быстро плавает при помощи ритмических сокращений своего «колокола» и при соприкосновении с нежными частями человеческой кожи производит сильное раздражение. Более мелкие размеры имеет медуза сарсия (около 2 см) и раткеа (около 0,5 см (р. 8, 9)). Не следует думать, что организмы планктона населяют обитаемую толщу морской воды равномерно: некоторые из них держатся в теплой поверхностной воде, другие в холодной глубинной; большинство предпочитает свет (в первую очередь, конечно, водоросли) но некоторые — темноту. Ребровиков — Pleurobranchia можно находить на поверхности моря лишь зимой и весной, пока вода не нагрелась; летом они скрываются на глубине, в слое воды с температурой 10°. Равным образом и ночесветки не переносят очень теплой воды и во вторую половину лета редко встречаются на поверхности. В итоге, соответственно различным наклонностям составляющих его организмов, планктон изменяет свой состав не только на различных глубинах, но и по временам года. Конечно, изучение планктона представляет выдающийся теоретический интерес; но может показаться странным, что не меньше и практическое его значение. В самом деле, незаметная глазу живая масса планктонных организмов играет огромную роль в круговороте веществ моря. Мириады микроскопических взвешенных в воде водорослей, подобно наземным растениям, строят органическое вещество своего тела из минеральных составных частей воды, служа вместе с тем пищей животным зоопланктона. Как растительный, так и животный планктон служат главной или исключительной пищей целому ряду рыб. имеющих промысловое значение, напр., хамса, некоторые виды сельдей; наконец, последние пожираются хищниками вроде скумбрии и судака, опять таки служащими предметом промысла. Таким образом оказывается, что микроскопические водоросли фитопланктона в значительной мере определяют собою промысловую продуктивность моря. В самом деле, планктон Черного моря количественно не богат, с чем гармонирует сравнительно малая продуктивность этого моря; с другой стороны, теплые мутные воды Азовского моря буквально насыщены фитопланктоном, объясняя отчасти его огромную продуктивность, к которой мы еще будем иметь случай вернуться. Переходим теперь к рыбному населению Черного моря, образующему основу его нектона. В Черном море нет крупных акул. Единственная акула, регулярно встречающаяся здесь, так наз., катран — Acanthias vulgaris (р. 53) обычно не превышает длиною 1 метра, хотя попадаются экземпляры свыше 16 кг весом. Подобно своим более крупным родичам катран — прожорливый хищник, иногда начисто выедающий из сетей весь улов рыбака. В пищу катран употребляется редко, хотя из него выходят недурные балыки. То же самое относится и к плоским родичам акул — скатам; в Черном море их водится два вида: гладкий скат или морской кот, обычно в 8—12 кг весом, живородящий (так же, как и катран) и снабженный на хвосте острою пилой, которой он может наносить опасные, иногда смертельные поранения. В Каркинитском заливе попадаются чудовищные коты до 98,4 кг весом. В Керченском проливе котов давно уже ловят для вытопки жира из их печени. Колючий скат или морская лисица (р. 57) лишен хвостовой пилы, размножается крупными яйцами, облеченными роговой скорлупой (р. 60), и не достигает таких размеров. Скаты — хищные рыбы, придерживающиеся ровного дна, и потому летом наиболее обычны в биоценозах песка и зостеры. Осетровые рыбы держатся в море круглый год, но половозрелые экземпляры уходят на летние месяцы в реки — Дон, Кубань, Днепр для икрометания. Зимой они возвращаются в Черное море и держатся на глубине 80—120 метров, где их ловят крючковой снастью, наживляемой мелкой рыбой, а в Керченском проливе и Азовском море «самоловными» крючьями. Севрюга — Acipenser stellatus ловится преимущественно в Азовском море, но также вместе с осетром (Acipenser Güldenstedti реже — A. sturio) у Евпаторийских и Феодосийских берегов; белуга — A. huso главным образом, по Ялтинскому побережью. Красная рыба, ловимая у крымских берегов, никогда не бывает крупной: средний вес осетра — 12 кг и не свыше 32 кг белуги 64—80 кг, редко больше 240 кг, севрюги — около 8 кг. Прежде уловы красной рыбы доходили до 100 тонн в год. Подобно осетровым и ловимые у берегов Крыма сельди (р. 51) относятся к проходным рыбам. Лов их производится, с одной стороны, в Керченском проливе и на побережьи от Керчи до Феодосии для сельдей, идущих метать икру в Азовское море и Дон (преим. Alosa pontica), с другой — на западном побережьи, кончая Севастополем, для видов, входящих в Днепр и Дунай — A. pontica. A. Nordmanni Ant. Главный сезон лова — осень, когда отъевшаяся за лето рыба входит в море, меньше — весной, при обратном ходе. Лов производится огромными неводами и сетями, в одной Керчи вылавливаются свыше 10 млн штук. Большое промысловое значение имеет также анчоус или хамса — Engraulis encrasicholus. Это — пелагическая, т. е. держащаяся в открытом море и питающаяся планктоном рыбка, летом «пасется» в мелководном Азовском море («азовская хамса») или в С.-З. углу Черного моря («черная хамса»), а зимой подходит к берегам Крыма неисчислимыми массами. Особенно много подходит к берегам Феодосии и Ялты азовской хамсы в зимы, когда преобладают восточные ветры. Как раз в самые голодные годы с 1919 по 1922 г. благодетельная хамса выручала население Крыма от голода, особенно обо-стрившегося весной. Ловят хамсу ночью наметами, а днем мелкоячейными неводами. В добычливые годы уловы бывают колоссальны и, конечно, превышают 1500 тонн. Кефаль (р. 59) тоже принадлежит к числу важнейших промысловых рыб Крыма и относится к нескольким видам, из коих наиболее крупный лобан — Mugil cephalus ловится реже; главную массу уловов составляют остронос — M. chelo и мелкая кефаль — M. auratus, M. saliens. Кефаль придерживается берегов, где питается органическими веществами ила и ловится здесь, в сущности, целый год. Ловят ее сетями, наметами, а также подъемными заводами — грандиозными сетяными сооружениями, подвешиваемыми к нескольким вышкам. Уловы кефали, поднимаемой одними заводами, достигали 700 тонн или 12 000 000 штук в год. Скумбрия (р. 50) тоже важная промысловая рыба; это типично-пелагический хищник, появляющийся у берегов Крыма в мае и уходящий в октябре в Мраморное море, где в марте месяце происходит его икрометание. Ловят скумбрию сетями и заводами, иногда «цапарями», т. е. удочками, закидываемыми с лодок на ходу. Добыча одних заводов достигала 3 млн штук. Огромный родич скумбрии — тунец — Thynnus thynnus, по местному мегала ставрида, достигающий двухсаженной длины и весящий десятки пудов, довольно часто появляется в Черном море, но местные рыбаки не умеют ловить его. Иногда попадается в заводы и меч-рыба — Xiphias gladius с мечевидно-удлиненной верхней челюстью, которой она поражает свою добычу и врагов. Рыбаки зовут ее «шпагой». Довольно большое промысловое значение имеет похожая на скумбрию сафритка — Trachurus trackurus, во множестве ловимая в начале зимы в Балаклаве. Мы уже упоминали о султанке и камбале, ведущих придонный образ жизни. Султанка — необычайно вкусная рыбка, ловимая преимущественно весной и осенью бочкообразными сетяными ловушками. Зимой она уходит на глубину. То же самое относится и к смариде или морскому окуньку — Smaris chryselis — красивой маленькой рыбке, ловимой вместе с султанкой мережками, но также и на удочку. Камбалы являются тоже важной промысловой рыбой. Широкоячейными сетями и наживными крючьями их вылавливают у берегов Крыма; во всяком случае десятки тысяч пудов. Из других морских рыб, имеющих лишь малое промысловое значение или вовсе не промысловых, упомянем следующих: лосось — Salmo trutta labrax в Черном море довольно мелок и у берегов Крыма попадается довольно редко. Гораздо чаще его ловят близ устьев бурных рек Кавказского побережья, куда он входит для икрометания. Сарган или морская щука, иначе «иголка» — Belone acus, змеевидная рыба с вытянутым в клюв рылом, иногда целыми стаями выскакивает из воды. Вкусным мясом саргана прежде пренебрегали из-за ярко-зеленого цвета его костей. Морской петух (р. 52) — красивая рыба с широкими грудными плавниками лазоревого цвета, высоко ценимая гастрономами. Лавраки или «белорыбица» — Morone labrax — крупная вкусная рыба, которую бьют ночью острогой рыбаки — «огневщики». Морской карась — Sargus annularis и каменный окунь, Serranus scriba замечательные тем, что они нормально являются гермафродитами, т. е. имеют и яичники и семенники. Звездочет или морская корова — Uranosropus scaber. безобразная рыба с направленными вверх глазами, неподвижно лежащая на дне. Морской чорт — Lophius piscatorius — отвратительное создание с необъятной зубатой пастью, приманивающее добычу червеобразным придатком 1-го луча спинного плавника. Мизгит или пикша — Gadus euxinus, мелкий родич северной трески, имеющий у нас лишь ничтожное промысловое значение и т. д. Кроме рыб, к нектону относятся дельфины, принадлежащие к китообразным млекопитающим. У берегов Крыма водится по меньшей мере три вида дельфинов. Самым обычным и вместе с тем наиболее жизнерадостным из них является белобочка — Delphinus delphis (р. 69), стада которого круглый год бороздят прибрежные воды Крыма в погоне за рыбой, которую эти зубатые хищники истребляют в огромном количестве. Белобочка достигает в среднем 82—98 кг весом. Более мелким видом является «пыхтун» — Phocaena relicta (р. 68), появляющийся у крымских берегов, главным образом, зимой, вместе с азовской хамсой. Вес 49—65 кг. Самый крупный, но и самый редкий дельфин офалина — Tursiops tursio, некоторые экземпляры которого достигают свыше 320 кг весом. Удивительная быстрота передвижения дельфинов: им ничего не стоит обогнать пароход, идущий 23 км в час. За последние годы дельфины служат предметом оживленного промысла. Обычно их бьют из ружей или винтовок с лодок и моторных катеров, главным образом, весной, когда их привлекает подходящая к берегам хамса. Раньше использовали для технических целей только их жир, образующий толстый подкожный слой так называемую, «шубу», весящую у белобочки и пыхтуна от 8 кг до 16 кг, а у офалины несколько десятков килограммов. Однако, минувшие голодные годы приучили население употреблять в пищу как жир, так и мясо дельфинов, и страшною зимою 1921—1922 г. сотни бедняков были спасены от голодной смерти дешевым и питательным, хотя и вонючим, мясом дельфина. Колбасные продукты из этого мяса одно время регулярно фигурировали на рынках приморских городов.
Типичные животные Черного моря (по С.А. Зернову). Объяснение рисунков. 23. Мидия, ракушник съедобный — Mytilus galloprovincialis L. 24. Морской гребешок — Pecten ponticus B. D. D. 25. Фолада — Pholas dactylus L. 26. Питалида, блюдечко — Patella pontica. Mil. 27. Солен, черенок — Solen vagina L. 28. Корабельный червь — Teredo navalis L. 29. Сверлящий моллюск петриколя — Petricola lithophaga Retz. 30. Нора петриколи (см. № 29). 31. Венус — Venus gallina L. 32. Устрица — Ostrea taurica Kryn. 33. Тапес — Tapes rugatus B. D. D. 34. Кардиум, сердцевидка — Cardium edule. 35. Артемия, соляная матка — Artemia salina Leach.; около l,5 сантиметра длиной, временами заполняет все соленые озера. 36. Рак-отшельник — Diogenes varians Hell. 37. Гульдия — Goiildia minima. 38. Морской желудь — Balanus improvisus Darw. 39. Ветвистоусый рачек подон — Podon 40. Морская коза — Caprella. 41. Мраморный краб — Pachygrapsus marmoratus Simps. 42. Насса — Nassa reticulata L. 43. Веслоногий рачек сем. — Copepoda. 44. Креветка — Leander. 45. Гаммарус, морская блоха — Orchestia. 46. Морской таракан — Idotea tricuspidata. 47. Травяной краб — Carcinus moenas L. 48. Ботриллюс — Botryllus schlosseri Sav. 49. Асцидия — оболочник — Ascidiella aspersa Müll. Примечание. Рисунки № № 39, 43 изображают собою животных, почти невидных невооруженным глазом В настоящее время тюлень — Monachus albiventer (рис. 71) окончательно истреблен у берегов Крыма и встречается лишь на противоположном Малоазиатском берегу Черного моря и берегов Болгарии. Но лет 50—60 тому назад тюленей еще видали на скалах около Херсонесского маяка и на Южном берегу. Азовское море, в общем, гораздо богаче рыбой, но состав его ихтиофауны совсем иной. Основу ее составляют рыбы солоновато-водные, вообще говоря держащиеся в предустьевых пространствах моря и лиманах: тарань, шемая, рыбец, чехонь, тюлька — Harengula cultriventris, атерина — Atherina pontica, сопач — Percarina maeotica, пухлощекая игла — Syngnathus bucculentus, глосса и множество видов бычков — Gobiidae. Весьма видную роль в рыбном населении играет также проходная рыба, подолгу кормящаяся в необычайно богатом пищей Азовском море: белуга, севрюга, осетр, шип — Acipenser schyp, сельдь. Из настоящих морских рыб сюда заходят сравнительно немногие: кефаль, хамса, султанка, сарган, камбала — Bothus maeoticus и некоторые другие. Взамен этого из лиманов и устьев рек в Азовское море заходит много настоящей пресноводной рыбы: шаран или карп, чебак или лещ, сом, щука, окунь, шерешпер, изредка стерлядь, и наконец, судак, за последние годы страшно размножившийся за счет пожираемых им бычков. Вследствие неумеренного вылова некогда неисчерпаемые рыбные богатства Азовского моря за последние десятилетия сильно растаяли. Некоторые породы, ловившиеся раньше в огромном количестве, как, напр., пузанок — Alosa tanaica и тарань — Rutilas rutilus Heckeli в настоящее время дают лишь незначительные уловы. Обратимся теперь к рассмотрению придонных организмов, или бентоса. Различные организмы этой биологической группировки распределены по дну моря в зависимости от глубины, характера берегов и грунта, образуя, так называемые, растительно-животные сообщества или биоценозы. Согласно исследованиям проф. С.А. Зернова, значительно дополненным и уточненным экспедициями последних лет, в Черном море можно различать следующие основные биоценозы. 1. Биоценоз скал, развитый на утесистых берегах от линии прибоя и до глубины, примерно 28 м. Этот биоценоз особенно пригоден для изучения прямо с берега, при помощи простого сачка или скребка. Линия прибоя отмечена поясом розоватой известковой водоросли кораллины; выше уровня воды шныряют по камням морские мокрицы — Lygia Brandtii Rathke; к скалам плотно присосались конические шапочки моллюска Patella (рис. 26) по местному «питалиди», употребляемые в пищу, и бесчисленные белые колпачки усоногих раков Balanus (рис. 38). В расщелинах скал сидят мелкие моллюски Littorina; местами скалы одеты целыми щетками двустворчатых моллюсков мидий (рис. 23), тоже съедобных. Все эти животные сидят очень плотно и обладают прочной скорлупой, иначе они были бы смыты и раздроблены прибоем. Ниже скалы покрыты густыми зарослями бурой водоросли Cystosira (рис. 10), которую рыбаки называют «бородой»; еще глубже из водорослей растут ярко-зеленые кусты морского салата (рис. 19), пузырчатой водоросли Enteromorpha (рис. 22) и багряной водоросли — Phyliophora (рис. 21). Если скалы достаточно мягки, как мергелистые утесы окрестностей Севастополя, они часто бывают источены сверлящими двустворчатыми моллюсками Pholas, Barnea и Petricola (рис. 25, 29), последний, впрочем, сверлит и твердые прибрежные известковые скалы Южного берега. Деревянные сооружения, равно как и обшивку парусных судов сверлит «корабельный червь» — тоже двустворчатый моллюск (рис. 28). На камнях, особенно под камнями, сидят «морские анемоны» или актинии (р. 18). Расщелины скал и заросли водорослей населены богатой фауной различных гидроидных полипов (р. 14), мшанок, образующих иногда целые наросты — Membranipora (р. 70), — губок — Halichondria (р. 17), Reniera, Spongelia, ярко желтых колониальных оболочников — Botryllus (рис. 48), немертин, креветок (рис. 44) и др. мелких рачков (рис. 40), моллюсков — Trochus и Nassa. По скалам шныряют бойкие черные крабы — Pachygrapsus marmoratus (р. 41), в расщелинах сидят крупные красные «каменные» крабы — Eriphia spinifrons с мощными клешнями, которых ночью часто ловят с огнем. Из рыб в скалах держатся мелкие морские собачки (рис. 61), больно кусающиеся, пестрые зеленушки, среди которых особенно эффектен необычайно ярко окрашенный Crenilabrus pavo (рис. 56), изящные черные «ласточки» или морские монахи — Heliastes chromis, безобразные морские ерши — Scorpaena porcus, вооруженные сильными колючками; угревидные галеи — Motella tricirrata. Все эти рыбы легко идут на удочку, на приманку из лапки краба или мяса мидии. 2. Биоценоз песка. Ровные песчаные пляжи встречаются, главным образом, на Евпаторийском и Феодосийском побережьях; на Южном берегу преобладают галечные пляжи, отличающиеся чрезвычайно бедной фауной. Настоящий же песчаный берег, уходящий вглубь метров на 28, содержит довольно богатое население. Преобладают различные роющие формы, с приостренным передним концом тела. Ближе к берегу скрываются в песке кольчатые черви Saccocirrus и Polygordius, немертины (рис. 11), глубже примитивное позвоночное животное ланцетник (рис. 67), в Крыму особенно многочисленный против Георгиевского монастыря, и маленькая рыбка Ammodytes, называемая рыбаками «емь». Из других рыб песчаное дно предпочитают бычки (рис. 62), неуклюжие рыбки с присоской на брюхе, плоские камбалы (рис. 63), султанка (рис. 66), разгребающая песок своими усиками, и так называемая змейка или морской скорпион (рис. 65), наносящий своими ядовитыми колючками весьма болезненные уколы. Чем глубже, тем многочисленнее становятся моллюски — вездесущая Nassa (рис. 42), сердцевидки (рис. 34), гребешок (рис. 24), венерка (рис. 31), гульдия (рис. 37). Под Керчью в песке много удлиненных «черенков» (рис. 27), употребляемых в пищу, кажется везде, кроме крымского побережья.10 Из раков в песке держится краб-плавунец Portunus, ловко плавающий расширенными задними лапками, и рак-отшельник (рис. 36), прячущийся в мертвую раковину какого-нибудь брюхоногого моллюска. 3. Биоценоз зостеры. В тех местах, где вследствие защищенности от прибоя напр., в бухтах), или благодаря выносу речками, к песку примешивается ил, фауна меняется; на глубине нескольких саженей развиваются подводные луга морской травы — Zostera marina, употребляемой в высушенном виде для набивки тюфяков. Эти заросли и образуют биоценоз зостеры. В иловатом грунте, под корнями зостеры роют свои ходы многочисленные черви (рис. 12, 13) и прячется единственная в Черном море морская звезда Amphiura (рис. 20); сама трава покрыта колониями гидроидов и мшанок и дает приют морским тараканам (рис. 46), креветкам (р. 44) и оригинальным рыбкам — морским иглам и морским конькам (рис. 55, 58), встречающимся, впрочем, и в цистозире. Тут же во множестве держатся «травяные крабы» — Carcinus maenas (рис. 47).
Типичные животные Черного моря (по С.А. Зернову). Объяснение рисунков. 50. Скумбрия, баламут — Scomber scomber L. 51. Сельд. сем. — Clupeidae. 52. Морской петух — Trigla hirundo B 1. 53. Акула катран или морская собака — Acanthias vulgaris Risso. 54. Белуга — Huso huso L. 55. Морская игла — Syngnathus sp. 56. Рулена, из зеленушек — Crenilabrus pavo Вrün. 57. Морская лисица, скат — Raja clavata L. 58. Морской конек — Hippocampus brevirostris Cuv. 59. Кефаль сем. — Mugilidae. 60. Яйцо ската, морской лисицы (рис. 57). 61. Морская собачка сем. — Blenniidae. 62. Бычек сем. — Gobiidae. 63. Камбала — Bothus maeoticus Pall. 64. Морской кот, скат — Trygon pastinaca Cuv. 65. Морской скорпион — Trachinus draco L. 66. Султанка или барбуня — Mullus barbatus L. 67. Амфиоксус (контур) — Amphioxus lancolatus Jarr 4. Биоценоз ракушечника. На глубине 27—37 метров песчаный грунт неизменно становится более плотным от примеси ила, ибо волнение становится все менее и менее чувствительным с глубиной; в связи с этим моллюски, особенно мидии и гребешки, становятся особенно многочисленными; местами среди них преобладают устрицы (рис. 32), которых добывают при помощи драг, и при посредстве которых только и возможно ознакомление с придонной фауной. Устриц прежде искусственно выращивали в особых заводах, напр., в Севастопольской бухте, добывая их по всему Крыму до 4 миллионов штук в год. На глубине 37—65 м грунт, вследствие преобладания ила, становится непригодным для устриц, и мы можем говорить об особом биоценозе мидиевого ила. 5. В биоценозе мидиевого ила из моллюсков преобладают мидии, однако, с гораздо более тонкими створками, чем в полосе прибоя, и модиолы — Modiola adriatica. Помимо этого, в биоценозах 4 и 5 живут круглые красные губки — Suberites (рис. 133), асцидии (рис. 49), глубоководные креветки — Crangon, иногда редкие в Черном море иглокожие — голотурии Cucumaria и Sinapta. Створки ракушек часто бывают покрыты известковыми трубками сидячих кольчатых червей (рис. 16). В илу держится интересный по своей анатомии червь Phoronis. Глубже 75 метров начинает преобладать мелкая ракушка Modiola phaseolina, створки которой покрыты волосками, и мы вступаем в область биоценоза фазеолинового ила. 6. Биоценоз фазеолинового ила покрывает обширные площади до глубины 146—180 м. Водоросли на этих мрачных, холодных глубинах почти отсутствуют, и животное население чрезвычайно бедно. Створки самой фазеолины проедаются хищными моллюсками Trophon, в иле сидят черви Phoronis, но главным образом — черви Terebellides carnea и Mellina adriatica, заключенные в трубки и являющиеся преобладающими формами этого биоценоза. Попадаются также амфиуры и голотурии и глубоководные креветки. Ниже 150 м, докуда доходит лишь один червь Mellina, мы обычно не встречаем уже никаких животных. Как и в случае пелагического населения (т.е. организмов открытого моря), зараженность глубин сероводородом кладет непреодолимый предел вертикальному заселению морского дна. Глубже всего спускается нижняя граница жизни у Кавказских берегов, где удалось доказать существование живых организмов с глубин свыше 200 метров. У берегов Крыма последние следы жизни прекращаются, уже начиная с глубины 130—140 метр.; вязкий вонючий черный ил, покрывающий окраины глубокой котловины Понта, помимо бактерий содержит лишь мертвые раковины моллюсков, некогда живших в Черном море, и микроскопические скелеты диатомовых водорослей, непрерывным дождем сыплющихся сверху из населенной толщи моря. Вытащенный на поверхность, этот ил быстро сереет вследствие окисления на воздухе содержащегося в нем гидрата односернистого железа. На самых больших глубинах цвет ила переходит в голубой или синий от большого содержания углекислой извести, наличие которой лишь в малой мере объясняется наличием известковых скелетов планктонных организмов, но она образуется, как мы видели, параллельно с сероводородом из растворенного в воде гипса. Таким образом, практически говоря, в Черном море заселена лишь, так называемая, континентальная ступень, в то время как вся, так называемая, абиссальная или глубоководная фауна, в Средиземном море и Атлантическом океане заключающая в себе целый мир разнообразных и причудливых животных, совершенно отсутствует в Черном море, глубокая котловина которого является настоящим царством смерти. Конечно, набросанная выше основная схема состава и распределения черноморских биоценозов подвержена многочисленным вариациям. Так, в заливах и закрытых бухтах илистые биоценозы всегда лежат на значительно меньшей глубине. Весьма замечателен биоценоз филлофорного моря, открытый С.А. Зерновым в С.-З. части Черного моря, в треугольнике Севастополь—Дунай—Одесса. Огромная площадь морского дна, на глубине около 60 м густо заросла красною водорослью филлофорой, заросли которой дают приют сравнительно небогатой фауне исключительно красных животных — рыб, червей, ракообразных.
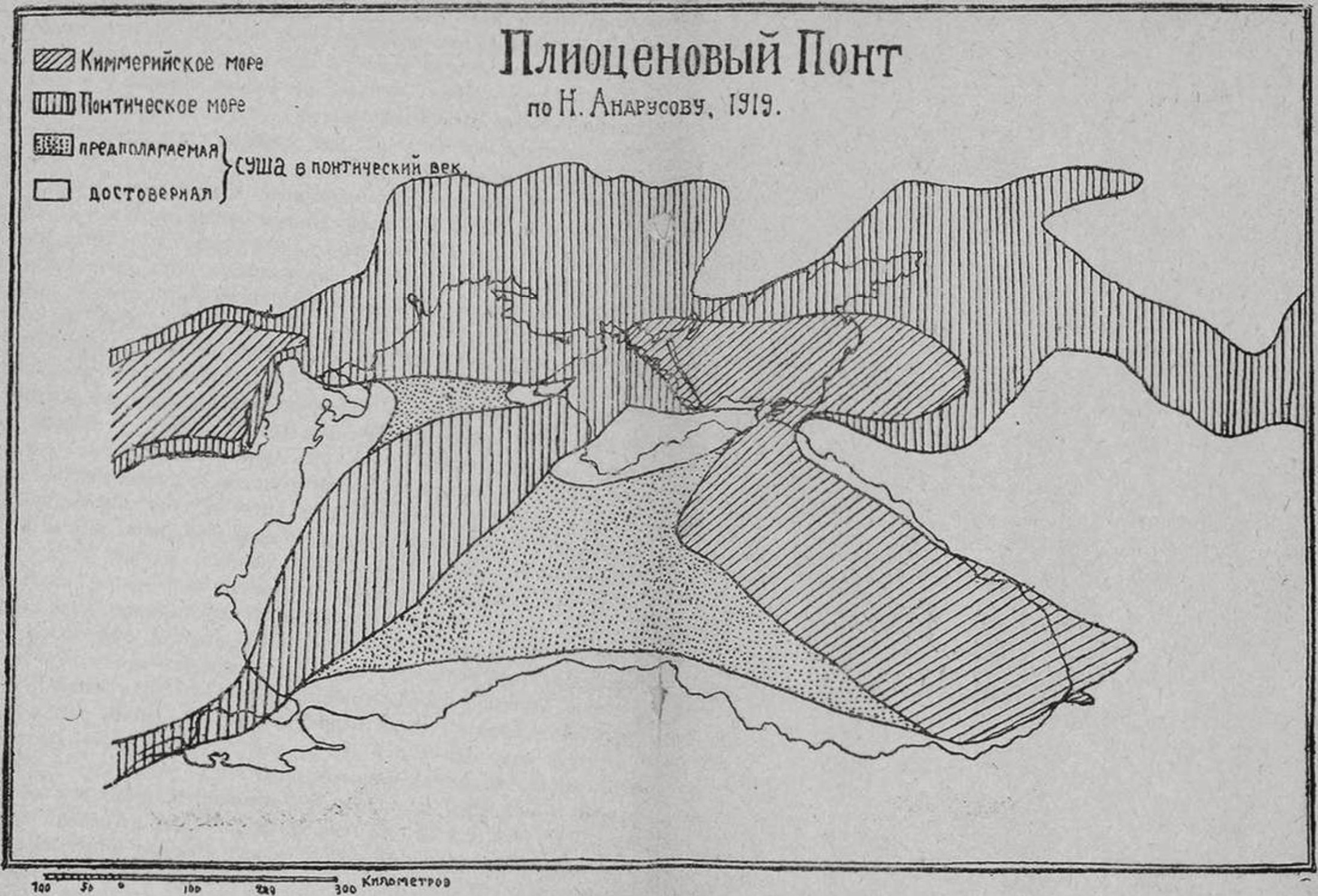
Придонное население Азовского моря, детально изученное за последние годы трудами Азовской экспедиции, сильно отличается от Черноморского, как и следовало ожидать, учитывая различия гидрологических условий. Скалистые берега развиты здесь лишь местами по северному побережью Керченского полуострова, вообще же берега Азовского моря опоясаны широкими пляжами преимущественно ракушечного песка. Биоценоз скал сразу же отличается от черноморского отсутствием зарослей цистозиры, встречающейся лишь в Керченском проливе; заменяющая ее красная водоросль кишит огромным количеством рачков — идотей, сфером и бокоплавов. Камни покрыты всюду мелкими баланусами, мидиями, но питалиды — Patella тоже отсутствуют. Среди камней снует множество водяных ужей, охотящихся на бычков. Биоценоз прибрежного песка простирается от уреза воды до глубины 1,5—3 м. Наиболее характерной формой является роющийся в песке рачек-бокоплав — Pontogammarus maeoticus, местами встречающийся в столь огромном количестве, что отпечаток ноги прошедшего в полосе прибоя человека моментально начинает кишеть массой потревоженных рачков. Прибрежные жители добывают их в любом количестве для корма уткам, просеивая песок сквозь решето. В песке встречаются отдельные моллюски — Cardium edule, Mytilaster Monterosatoi и др., черви — Nephthys. Биоценоз ракушечника опоясывает берега Азовского моря на глубине от 3 до 10 м. Благодаря тому, что песок уплотняется здесь некоторою примесью ила, моллюски, встречающиеся в предыдущем биоценозе единицами, здесь выступают массами. Опоясывающая море полоса ракушечника достигает большой ширины (до 16 миль) и покрывает, в общем, огромную площадь. Она отличается весьма густым населением, и потому высокою продуктивностью, являясь настоящим подводным пастбищем для целого ряда ценных промысловых рыб, особенно осетровых.11 Биоценоз илистого ракушечника соответствует в Азовском море мидиевому илу Черного и занимает полосу от 10,5 до 12,5 м глубиной. Незначительность глубины, на которой в Азовском море становится возможным отложение ила, объясняется огромным количеством мути, взвешенной в его воде и приносимой бурною Кубанью и могучим Доном.12 Фауна илистого ракушечника представляет из себя лишь сильно обедненное население верхнего ракушечника. Продуктивность этого биоценоза невелика. Биоценоз синдесмиевого ила. Вся центральная часть Азовского моря с глубиной 11—13 м занята биоценозом серого синдесмиевого или гидробиевого ила. Ил этот — полужидкой консистенции и нередко с явственным запахом сероводорода. Население его — чрезвычайно бедное и состоит из очень немногих видов: моллюск синдесмия и гидробия, червь Nephthys и похожая на ерша рыбка сопач Percarina maeotica. В самом Керченском проливе наблюдается постепенное обогащение Азовской фауны черноморскими видами. Такою же бедностью отличаются и некоторые части Таганрогского залива, где тоже наблюдается крайне изменчивый гидрологический режим. Таганрогский залив является настоящим лиманом реки Дона, и фауна его, как и вообще лиманов, слагается из трех элементов: ближе к выходу в море преобладают морские виды, в центре — специфически-лиманные, солоноватоводные, с примесью речных, а у устья Дона — настоящие речные, с примесью лиманных. Вышеупомянутая лиманная фауна, встречающаяся, помимо Таганрогского залива, также в Ахтанизовском и Сладковском лиманах Азовского моря, в Днепровско-Бугском, Днепровском и прочих лиманах Черного моря, представляет колоссальный теоретический интерес, благодаря своему большому сходству с фауной Каспийского моря. Из рыб в состав ее входят пуголовка, сопач, бычки, мелкая, похожая на бычка Gobiosoma caspium, колюшка, атерина, тюлька, из моллюсков — Dreissensia, Micromelania, Clessinia, различные виды сердцевидок с дегенерировавшим замком — Adanta, Monodacna. Как мы увидим дальше, состав этой фауны позволяет сделать ценные выводы относительно прошлого Черноморско-Азовского бассейна. Познакомившись в общих чертах с животным населением Черноморско-Азовского бассейна, попробуем проанализировать его особенности, по сравнению с фаунами других морей. Всякий, кому приходилось бывать на побережье Средиземного моря. Атлантического океана или даже на Мурманском берегу, неизменно испытывает разочарование при ознакомлении с фауной Черного моря — настолько она сера и однообразна, настолько в ней мало крупных, эффектных форм, присущих всякому «настоящему» морю. И в самом деле, в Черном море отсутствуют целые классы типичнейших морских животных; совершенно нет морских ежей, настоящих морских звезд — Asterioidea, кораллов (если не считать прибосфорского участка), банных губок, каракатиц и осьминогов. В планктоне отсутствуют радиолярии, яркие сифонофоры, киленогие и крылоногие моллюски, сальпы. Больше того: имеющиеся налицо типично-морские формы — голотурии, моллюски, нередко удивительно мелки и невзрачны. Между тем, за порогом Босфора, в Мраморном море (в глубоких его слоях), мы встречаем уже почти нормальную морскую фауну. В то время, как в соседнем Средиземном море насчитывается не менее 7000 видов многоклеточных животных, в Черном море число это вряд ли превысит тысячу. И все же, анализ видового состава фауны Черного моря показывает, что в ней явно преобладают виды, общие со Средиземным морем; по подсчету, произведенному С.А. Зерновым в 1912 году, оказывается, что из 801 известных в то время многоклеточных животных 680, т. е. около 78% встречаются и в Средиземном море. Как и следовало ожидать, чисто морские, т. е. атлантико-средиземноморские виды далеко неравномерно распределены в Черноморско-Азовском бассейне, правильным образом убывая по направлению от Босфора к Азовскому морю. Если обозначить число видов брюхоногих и двустворчатых моллюсков, обитающих в восточной части Средиземного моря через 100, то в Мраморном море из этого количества мы найдем лишь 84, в Босфоре 58—37, в Черном море — 22, и. наконец, в Азовском — всего 4—6. За вычетом атлантическо-средиземноморских видов остальные 201 или 22% черноморской фауны являются эндемиками, т. е. коренными обитателями Черноморско-Азовского бассейна. В первую очередь к ним относятся представители той солоноватоводной или лиманной фауны, с которой мы уже ознакомились выше. Мы указывали также на замечательный факт, что значительное число лиманных видов встречается также в полупресном Каспийском море. Конечно, многие из указанных особенностей Черноморской фауны легко объяснить современными физико-географическими особенностями Черного моря. Ясно, что средиземноморские виды могли проникнуть в Черное море только через Босфор, причем вполне акклиматизироваться смогли только наиболее выносливые из них, правда, выказывая в некоторых случаях признаки вырождения. Для проникновения других видов, даже целых классов животных, пониженная соленость, низкие температуры Черного моря и зараженность глубин его сероводородом составляют непреодолимое препятствие. Опыты с акклиматизацией животных Мраморного моря в аквариумах Севастопольской биологической станции показали, что животные эти гибнут У нас от разных причин: так, морские ежи уже через неделю дохнут от пониженной солености, морские же звезды великолепно живут летом, выдерживая по полугоду, но неизменно гибнут, как только температура зимой падает ниже 10°Ц. Однако, другие черты, отличающие фауну Азовско-Черноморского бассейна — так, обилие в нем видов северно-атлантического типа, в некоторых случаях даже отсутствующих в промежуточном Средиземном море, наличие в его придонном илу полуископаемых створок лиманных моллюсков, неожиданное присутствие черноморских видов в С.-З. углу Азовского моря, обилие в нашем морском бассейне эндемических видов, и особенно удивительное сходство его лиманной фауны с каспийской — все эти черты не могут быть объяснены современными гидрологическими условиями, и для их уразумения приходится прибегнуть к геологической истории не только Черноморско-Азовского бассейна, но и вообще южных морей. Всякому известно, что земная кора, особенно некоторые, наиболее податливые ее участки, находятся в состоянии непрерывного колебания — области, ныне занятые сушей, некогда были морем, и наоборот. В нашу задачу не входит изложение эволюции всех морских бассейнов, сменивших друг друга в течение земной истории на месте современного Понта. Нас должен интересовать, главным образом, момент, когда в этих бассейнах начинают проглядывать черты, характеризующие современное Черное море. Таким моментом была средина третичного периода. В эоцене, т. е. в самом начале третичного периода. Крымские горы омывались с севера заливом огромного тропического моря Тетис, по берегам которого жили крупные нуммулиты. В олигоцене залив этот сменился холодным морем с фауной северного характера (сельдь Melleta, устрица Ostrea cochlea), что указывает на связь с Атлантическим океаном, осуществлявшуюся через Нижне-Дунайскую низменность и Западную Европу. Этот бассейн имел вполне морской характер еще в нижнем миоцене (первый средиземноморский ярус, первый морской цикл), но уже в среднем миоцене (второй средиземноморский ярус) начинаются определенные следы опреснения и изоляции от океана. Опреснение это было уже вполне определенно выражено в так называемом Чокракском и Спаниодонтовых бассейнах, занимавших северную часть современного Черного моря, и в конце среднего миоцена сохраняется лишь очень мало чисто морских форм. В верхнем миоцене мы наблюдаем опять значительное наступание моря — второй морской цикл. Замкнутый Сарматский бассейн разливается на огромное расстояние — от Альп до подножья Тяньшаня, от донецкой кристаллической гряды на севере до Дарданелл на юге. Среди этой колоссальной водной площади Кавказ возвышался наподобие острова, а горный Крым представлял из себя направленный к северу полуостров обширной суши, лежавшей на юг. От Средиземного моря Сарматский бассейн отделялся обширной Эгейской сушей, занимавшей место современного Архипелага. Таким образом, северная часть Черного моря, Каспий и Арал входили в состав одного и того же огромного внутреннего моря. Богатая фауна Сарматского моря носит чрезвычайно своеобразный характер, указывающий на некоторое опреснение, и на полную изоляцию от океана. Характер глубоководных фаций отложений Сарматского моря с несомненностью указывает на то, что глубокие части его были заражены сероводородом и отличались такою же безжизненностью, как и глубокая котловина современного Черного моря. Характерны двустворчатые Mactra podolica, Mactra caspia, Tapes gregaria, образующие целые банки. На месте теперешнего Керченского полуострова колониальные организмы — мшанки — Membranipora lapidosa строили рифовые сооружения. напоминающие атоллы Тихого океана. Из рыб в Сарматском море уже встречались хамса — Engraulis encrasicholus, атеринка — Atherina Bronni, из китообразных — Cetotherium priscum, из сиреновых — Manatus maeoticus, (что указывает на тропический характер моря), из ластоногих — тюлень — Phoca Pontica и т. д. К концу миоцена заканчивается 2-й цикл развития Понто-Арало-Каспийского бассейна: Сарматское море начинает сокращаться в размерах и распадается на ряд мелких бассейнов с обедненной фауной (Херсонский ярус). Начало плиоцена знаменуется новым расширением моря (3-й морской цикл), которое получает название Мэотического бассейна: бассейн этот тянется неправильной полосой с изрезанными краями от Румынии (Паннонский бассейн), до восточного берега Каспия, покрывая своими водами степной Крым и Предкавказье. Фауна в нем — солоноватоводная, вообще говоря еще сарматского типа. Конец 3-го морского цикла знаменуется сильнейшим сокращением Мэотического бассейна, в котором получают преобладание малорослые полупресноводные и пресноводные виды.
68. Дельфин — пыхтун. 69. Дельфин — белобочка. 70. Свая, обросшая мшанкой Membranipora. 71. Белобрюхий тюлень Мы подходим к весьма значительной и важной для нас фазе эволюции нашего морского бассейна. Снова широко разливаются воды моря (4-й морской цикл). Обширный Понтический бассейн простирается на север до кристаллической гряды, где древний берег его еще теперь можно проследить, следуя по изогипсе 120 метров над уровнем моря. На запад оно покрывает Средне-Дунайскую низменность, образуя Дакийский залив, посреди которого возвышаются два острова, на юг дает рукав во Фракию. Крым по-прежнему выдается в виде северного полуострова Понтической или Киммерийской суши, отделяясь от Кавказа широким и глубоким Таманским проливом. На востоке Понтическое море соединяется с Каспием и Аралом через Манычскую впадину; быть может, был пролив и через Закавказье, так что Кавказ мог и в Понтическое уже время быть островом. Глубина моря, за исключением Таманского пролива, Дакийского и Сухумского заливов, была незначительна, соленость — сильно понижена, вероятно, напоминая соленость современного Каспия. По-видимому, имело место и заражение глубин сероводородом. Фауна его составилась в своей основе из выходцев Паннонского бассейна (см. выше). Рыбы, населявшие Понтическое море, в значительной мере принадлежали к видам и поныне держащимся в лиманах, воды его бороздили дельфины — Phocaena relicta, и поныне живущие в Черном и Азовском морях. В средине плиоцена (Киммерийский век) условия жизни в Понтическом море сильно изменяются: снова начинается отступание моря, и распадение его на отдельные бассейны. Понтический бассейн надолго отделяется от Каспийского, отделяется от него и Дакийский залив, вода которого все более и более опресняется. Древний берег Понтического моря сильно отступает к югу, и Горный Крым отделяется от Украины лишь неглубоким морским проливом километров в 100 шириной. С этого момента мы уже не имеем уверенности в существовании на юг от Крыма Понтийской суши. Возможно, что резкое отступание моря на севере происходило вследствие начавшегося опускания Понтиды и образования глубокой впадины современного Черного моря, в которую должны были устремиться воды с севера (так называемое, эвстатическое отступание моря). Одновременно с вышеописанными пертурбациями в распределении суши и моря имели место и значительные изменения климата, который в Киммерийском веке имел тропический характер, по крайней мере у Керченских и Кавказских берегов, где отлагались бурые железняки в результате размывания красных латеритовых почв Кавказа.13 В южной части Киммерийского бассейна обитали крупные, оригинальные моллюски совершенно тропического (полинезийского) типа. В верхнем плиоцене снова начинается некоторое расширение моря, но климат стал меняться в худшую сторону, на что указывает обеднение и измельчание фауны моллюсков в продолжение Куяльницкого и последовавшего за ним Чаудинского веков. Большой интерес представляет фауна слоев, слагающих мыс Чауда близ Феодосии, найденная также у Трапезунда и у Галлиполи, на юге Мраморного моря. Фауна эта состоит из видов, указывающих на замкнутый бассейн с пониженной соленостью и на захолодание климата; это заставляет, во-первых, предполагать, что Чаудинский век по времени совпадал с первым, так называемым, Гюнцским оледенением, которое современные геологи вынуждены отодвинуть к концу плиоцена; во-вторых, нахождение Чаудинской фауны на юге Мраморного моря определенно указывает на соединение Эвксинского бассейна с Мраморным морем, происшедшее благодаря опусканию в области Босфора, который представлял из себя раньше реку, текшую с Эгейской суши в Эвксинский бассейн, т. е. в Черное море. По-видимому, в конце плиоцена продвижение Эвксинского бассейна с севера на юг закончилось; согласно мнению некоторых исследователей (Андрусов, Hoernes и др.) в Чаудинский век (или непосредственно за ним) северный берег тогдашнего Черного моря значительно отступил к югу. Уровень Черного моря должен был быть метров на 30—50 ниже современного, и берег проходил в виде выпуклой к северу линии от мыса Эмине к южному берегу Крыма, а оттуда — к Новороссийску. Азовское море еще не существовало, а представляло из себя болотистую страну, прорезанную мощной рекой Доном и его притоками. Геологи по разному объясняют это продвижение береговой линии на юг: по Соколову — оно имело место благодаря «усыханию» моря, по Андрусову — благодаря поднятию суши, (так называемое, эпейрогенетическое движение земной коры). Не отрицая существования последнего фактора, мы все же должны согласиться с тем, что в замкнутом морском бассейне, каковым был верхнеплиоценовый Понт, опускание значительных масс суши неминуемо должно было вызвать отлив мелких вод на севере для заполнения все более и более углубляющейся и расширяющейся впадины на юге. Циклы отступания и наступания моря, описанные нами для Третичного периода, продолжаются и в Четвертичном, который, вообще, характеризуется многократными эпейрогенетическнми, идущими параллельно с наступаниями и отступаниями ледников, движениями14 земной коры. Принято думать, что периодам наступания ледников соответствует опускание суши, а периодам межледниковым — ее поднятие. Описанное выше низкое стояние уровня Черного моря, вероятно, соответствовало по времени 1-й (Гюнц-миндельской) межледниковой эпохе, которая и в других местностях Европы характеризуется как время наивысшего поднятия суши. За этим периодом сокращения моря последовала мощная его, так называемая, древнекаспийская или Хозарская трансгрессия. Снова восстановилась связь Черного моря с Каспием, фауна которого появляется на всем протяжении сильно опресненного Эвксинского бассейна, от Таганрога на востоке до Никополя и Джурджулет на западе. По миновании этой трансгрессии, вероятно, в наиболее теплый Рисс-Вюрмский межледниковый период наступает знаменательная страница в эволюции нашего морского бассейна: впервые после своего отделения от океана в средне-третичное время, оно приходит в связь с открытым морем, на этот раз — с недавно возникнувшим Эгейским. Благодаря опусканию суши в области Дарданелл, теплые и соленые Эгейские воды устремляются в Черное море, вытесняют каспийскую фауну и несут с собой совершенно новый мир животных южного моря. В береговых отложениях этого времени, которые сохранились в виде приподнятых террас во многих местах — близ Керчи, Тамани, Судака, Кавказского и Малоазиатского побережий, обнаруживается фауна, которая во многих отношениях приближается к современной черноморской, отличаясь от нее великорослостью и присутствием в ней до 14 видов моллюсков, отсутствующих в современном Черном море; некоторые из них в настоящее время совсем вымерли — Tapes Calverti и др., другие же — Cardium tuberculatum, Area None, Venus verrucosa и т. д. и теперь еще живут в Средиземном и Мраморном морях. Эта оригинальная фауна свидетельствует, во-первых, о более теплом климате, которым, вообще, отличались межледниковые периоды, а, во-вторых, о повышенной солености, которая, по мнению Н.И. Андрусона, должна была превышать 2%, т. е. соответствовала солености современного Мраморного моря. Впоследствии соединение Эвксинского бассейна со Средиземным морем было опять нарушено, и когда с наступлением последнего (Вюрмского) оледенения увеличение количества осадков, сопровождаемое, вероятно, понижением суши на север от Кавказа вызвало новую Каспийскую (Хвалынскую) трансгрессию холодного опресненного моря, фауна последнего, состоящая из тех же дрейссензий, микромеланий и беззамковых кардид вновь распространилась по всему протяжению не только Черного, но и Мраморного морей. Об этом красноречиво свидетельствуют мертвые, «полуископаемые» створки их, которые драга до сих пор выносит со дна этих морей, с глубины 100—275 м.
Ископаемый моллюск — Dreissensia rostriformis из глубинных осадков Черного моря Мы подходим к концу богатой событиями истории нашего морского бассейна. По окончании Вюрмского оледенения, в начале послеледниковой эпохи наступает, наконец, то событие, которое превратило его в современное Черное море. Вероятно, благодаря новому опусканию в области Дарданелл восстанавливается сообщение Черного моря со Средиземным, воды которого вторично вторгаются в Черное море.15 Благодаря происшедшему повышению уровня моря затопляются нижние части глубоких долин южнорусских рек — Буга, Днепра, Дона, в результате чего возникают обширные предустьевые озера — лиманы, каковыми по своему происхождению должны считаться также Севастопольская бухта и «Золотой Рог» Босфорского пролива. С другой стороны, соединение Азовского моря с Каспием через Манычскую впадину к концу послеледниковой эпохи прекращается. Теплые и соленые воды Средиземного моря вновь оттеснили полупресноводное население Черного моря в лиманы и Азовское море, где она существует и по сию пору в виде, так называемой, остаточной или «реликтовой» фауны. Меньшая часть составляющих ее видов, однако, сумела приспособиться к новому гидрологическому режиму, и показывает более широкое распространение (напр., бычки, сельди). Но главную часть фауны Черного моря составили все же средиземноморские переселенцы, которые нашли в себе достаточно пластичности и неприхотливости, чтобы приспособиться к жизни в сравнительно холодном опресненном бассейне. Мы видели, таким образом, что набросанная нами в общих чертах картина эволюции Черноморско-Азовского бассейна в достаточной степени объясняет нам казавшиеся раньше загадочными факты современного распространения организмов. Нам понятно присутствие в придонном илу Черного, Азовского и даже Мраморного морей створок полупресноводных раковин, населявших весь этот бассейн до прорыва Дарданелл, во время «Хвалынском» трансгрессии. Не требует пояснения и неоднократно отмеченное сходство лиманной фауны Черноморско-Азовского бассейна с современной Каспийской. Процесс заселения Черного моря средиземноморскими пришельцами, начавшийся после прорыва Дарданелл и определивший современную физиономию Черноморской фауны, несомненно еще не закончился: ряд видов рыб еще не может считаться окончательно укоренившимся в Черном море, так как придерживается южного и юго-западного берегов его («морской юнкер», сардинка); некоторые рыбы даже уходят для икрометания в Мраморное море (скумбрия, паламида). Единичные случаи нахождения даже у берегов Крыма омаров и других необычных для Черного моря беспозвоночных тоже как-будто намекают на то, что возможности колонизации и приспособления еще не исчерпаны, и фауна его со временем может обогатиться новыми пришельцами; впрочем, надо сознаться, что гидрологические условия Черного моря, даже помимо его общей пониженной солености, очень мало благоприятствуют такой иммиграции; путь через зараженные глубины закрыт, путь по западному берегу сильно затруднен огромными массами пресной воды, вливаемыми могучим Дунаем. Неудивительно, что некоторые рыбы, напр., сардинка, обычно и не идут севернее Румынии. Однако, у нас нет никакой уверенности в том, что установившийся в Черном море режим представляет из себя нечто стойкое, незыблемое. Мы видели, что на протяжении третичного и ледникового периодов морской бассейн, находившийся на месте Черного моря, то широко разливался, то, наоборот, сокращался, наконец, даже как бы передвинулся с севера на юг благодаря опусканию Понтийской суши; вся эволюция его представляется в виде гигантских пульсаций, и мы не имеем ни малейшей уверенности в том, что пульсации эти закончились. Ярким доказательством сказанного служит землетрясение, потрясшее в 1927 г. берега Черного моря и особенно сильно ощущавшееся в Крыму. В результате последних экспедиций, предпринятых Гидрографическим управлением и Севастопольской биостанцией, выяснилось, что южное побережье Крыма от меридиана мыса Тарханкут и примерно до меридиана Феодосии опоясывается на глубине около 1500 метров узкой полосой, где морское дно обнажено от ила; согласно показаниям сейсмографов, эта полоса и является эпифокальной областью последнего, еще не вполне закончившегося землетрясения, вдоль которой, вероятно, продолжается оседание дна Черного моря. Для всякого, знакомого с его историей, ясно, что последнее землетрясение является лишь затухающим отзвуком событий, начавшихся еще в конце третичного периода и совершавшихся в грандиозном масштабе в течение ледникового. Частые и мощные землетрясения, происходящие в области Мраморного моря, в среднем, каждые 50 лет, указывают на то, что земная кора находится там в состоянии еще более неустойчивого равновесия, чем у берега Крыма. Понижение суши в северо-западной оконечности Малой Азии (Скутарийский вилайет) на 40 метров не только бы углубило и расширило Босфор, но образовало бы новый пролив из Черного моря в Мраморное, через долину р. Сакарий, озеро Сабанджа и Измидский залив. Результатом этого несомненно явилось бы значительное осолонение и потепление вод Черного моря, параллельно с соответствующим изменением фауны. Наоборот, даже небольшое поднятие суши в области Босфора, по мнению некоторых ученых даже засорение его фарватера, снова приблизило бы Черное море к типу солоноватоводного бассейна с фауной Азовско-Каспийского типа. Доисторические обитатели крымских пещер, несомненно, уже были свидетелями грандиозных перемен в режиме и размерах Черного моря; не подлежит сомнению то, что и грядущему человечеству суждено пережить подобные пертурбации, быть может, даже вмешаться в них при помощи высоко развитой техники. В заключение нашего очерка нам хотелось бы подчеркнуть необычайно высокий интерес, представляемый изучением Черного моря, и его на первый взгляд бедной и невзрачной фауны и флоры. Конечно, ученый или дилетант, которого в фауне моря привлекает прежде всего величина, разнообразие и эффектность составляющих ее организмов, ошибется, попав на берега Черного моря; но исследователь, находящий удовлетворение в разрешении сложных и необычайно интересных зоогеографических и общебиологических проблем, найдет себе благодарную почву при изучении неказистой на первый взгляд фауны Черноморско-Азовского бассейна. Исследование морей и океанов, в частности мрачных глубин Эвксинского Понта, являет нам яркий пример мощи испытующего ума человека, который измерил море сначала только «очами жадными», а потом и во всеоружии точнейших физико-химических приборов и поистине проверил перед лицом моря силы своего духа. Проф. И. Пузанов. ЛитератураОстроумов Л.А. — Жизнь Южно-русских морей. Казань. 1902. Зернов С.А. и Андрусов Н.И. — Черное море. Статья в 1-м издании настоящего «Путеводителя». Симферополь. 1914. Совинский В.К. — Введение в изучении фауны Понто-Каспийско-Аральского морского бассейна. Записки Киевск. о-ва естествоиспытателей, т. XVIII. 1904. Скворцов Е.Ф. и Никитин В.Н. — Гидрологические разрезы, произведенные на Черном море в августе и ноябре 1923 года и в марте 1924 года. Записки по гидрографии, т. 49. 1925. Никитин В.Н. — Гидробиологические исследования Черного моря. Записки Крымского о-ва естествоиспытателей и любит. природы, т. VIII. 1925. Данильченко П.Т. и Чигирин Н.И. — К вопросу о происхождении сероводорода в Черном море. Труды особой зоологической лаборатории Академии Наук. Сер. II, №№ 5—10. 1926. Труды Азовско-Черноморской научно-промысловой экспедиции. Керчь. 1926. (Статьи: Книповича Н.М., Недошивина А.Я., Чугунова Н.Л.). Никитин В.Н. — Вертикальное распределение планктона в Черном море. Труды Особой зоол. лабор. Акад. Наук. Сер. II, 5—10. 1926. Зернов С.А. — К вопросу об изучении жизни Черного моря. Зап. Акад. Наук. Т. XXXII. 1913. Андрусов Н.И. — Взаимоотношения Эвксинского и Каспийского бассейнов в Неогеновую эпоху. Изв. Акад. Наук, т. 48. 1918. Андрусов Н.И. — Геологическое строение дна Керченского пролива. Изв. Акад. Наук. 1918. Андрусов Н.И. — Послетретичная Тирренская терраса. Прага. 1925. Оствальд Фр. Ф. — К истории тектонического развития Армянского нагорья. Зап. Кавк. отд. русск. геогр. о-ва, XXIX. 1916. Двойченко П.A. — Геологическая история Крыма. Зап-Крымск. о-ва ест., т. VIII. 1925. Скворцов Е.Ф. — Некоторые результаты Экспедиции по исследованию грунта Черного моря в связи с землетрясением. — Сборник «Землетрясения 1927 г. и судьбы Крыма». Симферополь. 1928. Архангельский А.А. — Об осадках Черного моря и их значении в познании осадочных горных пород. Бюл. Моск. о-ва испытателей природы. Отд. геолог., т. V. 1927. Примечания1. Управление безопасности кораблевождения Черного и Азовского морей. 2. Для сравнения приводим наибольшие глубины некоторых морских бассейнов: Балтийское море 427 м, Средиземное море 4400 м. Тихий океан 10 793 м (близ Филиппинских островов). 3. В океане наблюдались волны высотою в 13 метров. 4. Насчет этого ежегодного и вполне нормального понижения уровня следует отнести неоднократно появлявшиеся в прессе сенсационные известия об изменении уровня у крымских берегов, происшедшего якобы в результате землетрясения 1927 года. 5. Еще более горькой является вода Каспийского моря, где в сухом остатке соли мы находим 23,64% сернокислого магния и 6,99% гипса. 6. Абсолютный удельный вес или плотность воды равна в Черном море, в среднем, 1,01429—1,01123 на поверхности, 1,01631 на глубине 150 метров и 1,01735 на глубине 2000 метров. Плотность воды в океане равна при температуре 0°Ц 1,02813. 7. В Каспийском море придонная вода содержит 0,3—0,4 см³ сероводорода на литр. 8. Восстановление это, в случае, напр. сернокислый магнезии, происходит в 2 фазы, по схеме: I MgSO4 + С2 (opг. вещество) → MgS + 2 CO2; II MgS + H2O + 2 CO2 = H2S + Mg (HCO3)2. Правильность этой схемы доказывается тем, что с глубиной количество карбонатов, т. е. углекислых солей (в нашем случае Mg (HCO3)2 возрастает рука об руку с возрастанием количества сероводорода. 9. Таким образом, толща воды Черного моря разделяется на 2 химически противоположных зоны: поверхностную зону окисления, где доминируют окислительные процессы, и глубокую зону восстановления. Не следует думать, что процессы восстановления и окисления ограничиваются одними лишь сернокислыми соединениями: азот тоже восстанавливается из нитратов или азотнокислых солей в виде аммиака, или свободного азота, по схеме: NO3 → NO2 → NH3 → N. 10. Особенно распространен промысел на них у берегов Болгарии. 11. Продуктивность биоценоза определяется «сырым весом», или же числом индивидов животных, населяющих один квадратный метр грунта. Исследование этой продуктивности, имеющее огромное промысловое значение, сделалось возможным благодаря применению особых дночерпателей или храпов, захватывающих все население определенной площади. В общем, по своей продуктивности Азовское море стоит неизмеримо выше Черного. Максимальною продуктивностью отличается рассмотренный выше биоценоз ракушечника, в котором сырой вес организмов равен приблизительно 1 кило на квадратный метр, причем количество одного Cardium доходит до 1840 штук. Продукция в С.-В. углу Черного моря на глубине около 40 метров (ракушечник) равна приблизительно 140 гр. на кв. метр. 12. Вспомним, что в Черном море, вода которого содержит мало илистых частиц, иловатый грунт появляется лишь на глубине около 50 метров. 13. Латериты — красные почвы, образующиеся путем выветривания изверженных пород в условиях тропического климата. 14. Т.е. вертикальные поднятия и опускания больших площадей. 15. Согласно последним исследованиям акад. А.Д. Архангельского, изучавшего образцы ила, добытые последними Черноморскими экспедициями Гидрографического управления и Севастопольской биостанции, это событие должно было произойти даже не далее как 4000 лет тому назад, на что указывает число слоев тонкой пленки современного ила, покрывающего более глубокие слои ила с остатками полупресноводных моллюсков. Однако, эти данные нуждаются еще в подтверждении.
|
Столица: Симферополь
Крупнейшие города: Севастополь, Симферополь, Керчь, Евпатория, Ялта
Территория: 26,2 тыс. км2
Население: 1 977 000 (2005)
Крупнейшие города: Севастополь, Симферополь, Керчь, Евпатория, Ялта
Территория: 26,2 тыс. км2
Население: 1 977 000 (2005)

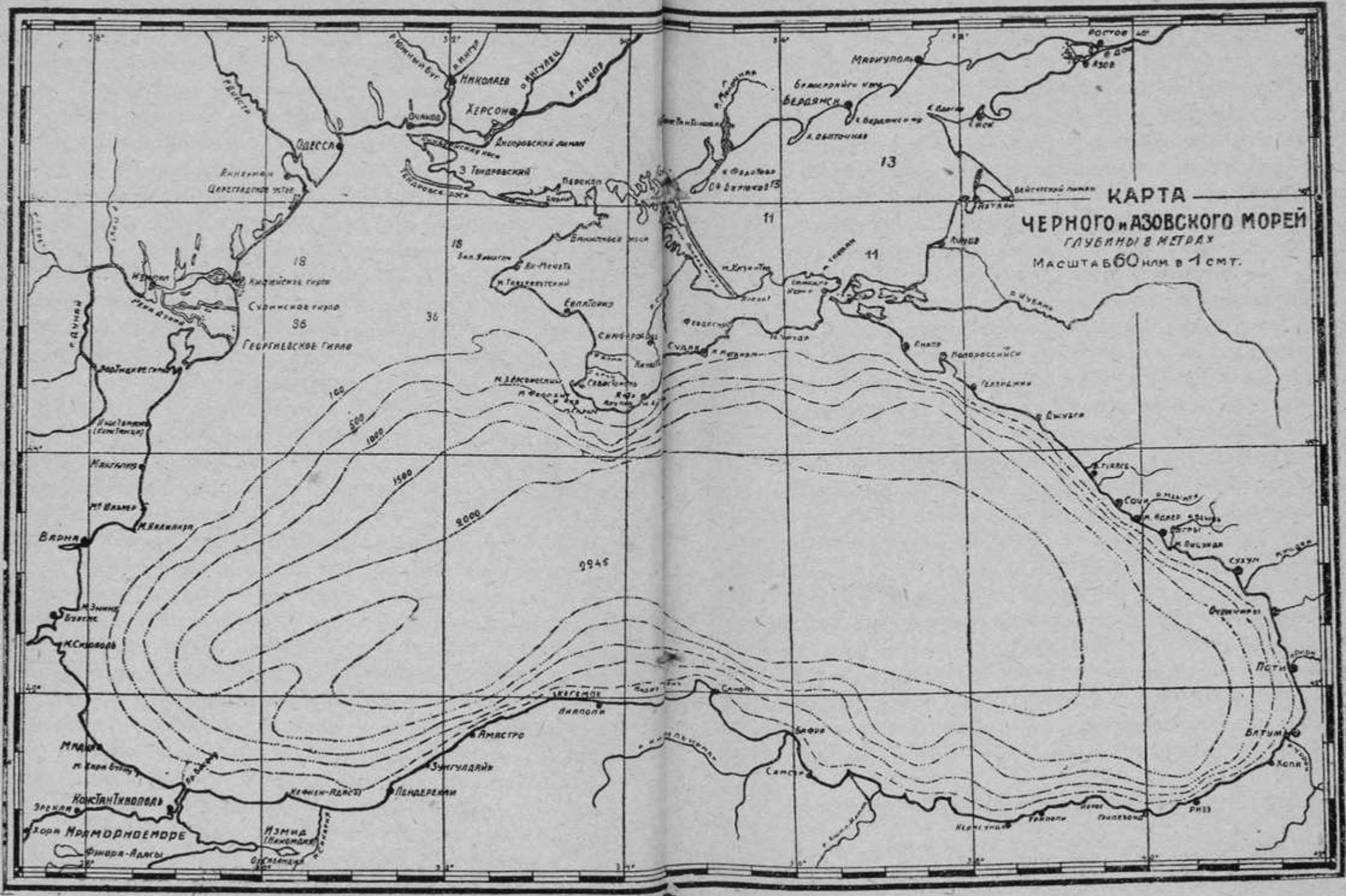
. Объяснение рисунков. 1. Медуза корнерот — Pilema pulmo Haeck. 2. Планктонная диатомовая водоросль хэтоцерос — Chaetoceros. 3. Медуза ушастая аврелия — Aurelia aurita M. Edw. 4. Ночесвет или ноктилюка — Noctihica miliaris Ehrb. 5. Перидиниевая водоросль церациум — Ceratium tripos. 6. Ктенофора или ребровик плейробранхия — Pleurobranchia pileus Fabr. 7. Перидиниевая водоросль гониавлакс — Gonyaulax polyedra Stein. 8. Медуза раткеа — Rathkea blumenbachii Rathke. 9. Медуза сарсия — Sarsia tubulosa Forbs. 10. Бурая водоросль цистозира — Cystoseira barbata L. 11. Немертина — Eunemertes gracilis Johnst. 12. Червь нереида — Nereis diversicolor Müll. 13. Червь пескожил — Arenicola grubii Clap. 14. Гидроид аглаофения — Aglaоphenia pluma Lmc. 15. Губка суберитес — Suberites domuncula, Schmidt. 16. Трубчатый червь — Potamoceros triquetroides Panc. 17. Кремневая губка халихондрия — Halichondria grossa Schmidt. 18. Актиния, морской анемон — Actinia equina L. 19. Зеленая водоросль морской салат — Ulva lactuca L. 20. Амфиура — Amphiura florifera Frbs. 21. Красная водоросль — Phyllophora rubens Grev. 22. Зеленая водоросль энтероморфа — Enteromorpha. Примечание. Рисунки № № 2, 4, 5, 7 изображают собою животных, почти невидимых невооруженным глазом")

. Объяснение рисунков. 23. Мидия, ракушник съедобный — Mytilus galloprovincialis L. 24. Морской гребешок — Pecten ponticus B. D. D. 25. Фолада — Pholas dactylus L. 26. Питалида, блюдечко — Patella pontica. Mil. 27. Солен, черенок — Solen vagina L. 28. Корабельный червь — Teredo navalis L. 29. Сверлящий моллюск петриколя — Petricola lithophaga Retz. 30. Нора петриколи (см. № 29). 31. Венус — Venus gallina L. 32. Устрица — Ostrea taurica Kryn. 33. Тапес — Tapes rugatus B. D. D. 34. Кардиум, сердцевидка — Cardium edule. 35. Артемия, соляная матка — Artemia salina Leach.; около l,5 сантиметра длиной, временами заполняет все соленые озера. 36. Рак-отшельник — Diogenes varians Hell. 37. Гульдия — Goiildia minima. 38. Морской желудь — Balanus improvisus Darw. 39. Ветвистоусый рачек подон — Podon 40. Морская коза — Caprella. 41. Мраморный краб — Pachygrapsus marmoratus Simps. 42. Насса — Nassa reticulata L. 43. Веслоногий рачек сем. — Copepoda. 44. Креветка — Leander. 45. Гаммарус, морская блоха — Orchestia. 46. Морской таракан — Idotea tricuspidata. 47. Травяной краб — Carcinus moenas L. 48. Ботриллюс — Botryllus schlosseri Sav. 49. Асцидия — оболочник — Ascidiella aspersa Müll. Примечание. Рисунки № № 39, 43 изображают собою животных, почти невидных невооруженным глазом")
. Объяснение рисунков. 50. Скумбрия, баламут — Scomber scomber L. 51. Сельд. сем. — Clupeidae. 52. Морской петух — Trigla hirundo B 1. 53. Акула катран или морская собака — Acanthias vulgaris Risso. 54. Белуга — Huso huso L. 55. Морская игла — Syngnathus sp. 56. Рулена, из зеленушек — Crenilabrus pavo Вrün. 57. Морская лисица, скат — Raja clavata L. 58. Морской конек — Hippocampus brevirostris Cuv. 59. Кефаль сем. — Mugilidae. 60. Яйцо ската, морской лисицы (рис. 57). 61. Морская собачка сем. — Blenniidae. 62. Бычек сем. — Gobiidae. 63. Камбала — Bothus maeoticus Pall. 64. Морской кот, скат — Trygon pastinaca Cuv. 65. Морской скорпион — Trachinus draco L. 66. Султанка или барбуня — Mullus barbatus L. 67. Амфиоксус (контур) — Amphioxus lancolatus Jarr")